![]()
![]()
![]()
![]()
Privatdozentin
Raum:
ND 2/136
Tel.: 0234 32 -
23830
Akademische Rätin
ORCID-iD: 0000-0001-8879-3348
E-Mail

Pro- und eukaryotische photosynthetisch aktive Mikroorganismen spielen eine enorm wichtige Rolle für das Ökosystem Erde. Sie sind für einen großen Teil der photosynthetischen Bildung von O2 und Biomasse und parallel der Fixierung von CO2 verantwortlich und sichern damit das Überleben der nicht-photosynthetischen Lebewesen einschließlich der Menschen. In ihrer natürlichen Umgeben erfahren alle Organismen häufig Stress, etwa durch den Mangel oder den Überschuss eines Nährstoffs. Photosynthetische Organismen müssen aufgrund der hochenergetischen Vorgänge der Lichtreaktionen besonders in der Lage sein, auf Stress mit geeigneten Abwehrmaßnahmen zu reagieren. Photosynthetische H2-Produktion durch Mikroalgen stellt solch eine Stressreaktion dar und dient der Ableitung überschüssiger Lichtenergie, um oxidative Schäden der Zelle zu vermeiden. Sie wird insbesondere bei Nährstoffmangel induziert, und dieser geht in Algenkulturen häufig mit der Etablierung O2-armer (hypoxischer) Bedingungen einher. Unsere frühen Forschungsarbeiten haben dazu beigetragen, den hoch-komplexen – und für einen Eukaryoten ungewöhnlichen – Gärungsstoffwechsel der Mikroalge Chlamydomonas reinhardtii zu verstehen, von welchem die H2-Bildung einen Teil darstellt. Neben faszinierenden Einblicken in die Ökologie dieser Alge, welche wiederum für funktionierende Ökosysteme ausschlaggebend ist, ist die Erforschung der Anpassungen von Mikroalgen an O2-Mangel auch biotechnologisch relevant. Zum einen stellen einige der Gärungsprodukte (z.B. Ethanol oder Milchsäure) industriell relevante Stoffe dar, zum anderen erfahren Algenkulturen in wirtschaftlich bereits etablierten Kultivierungsbecken häufig hypoxische Bedingungen. Neben der Charaterisierung des Stoffwechsels unter diesen Bedingungen versuchen wir auch zu verstehen, wie die Alge überhaupt ‚merkt‘, dass O2 fehlt, und wie sie diese Information in der Zelle weiterleitet, um die entsprechenden Antworten – oft auf Ebene der Genexpression – zu aktivieren.
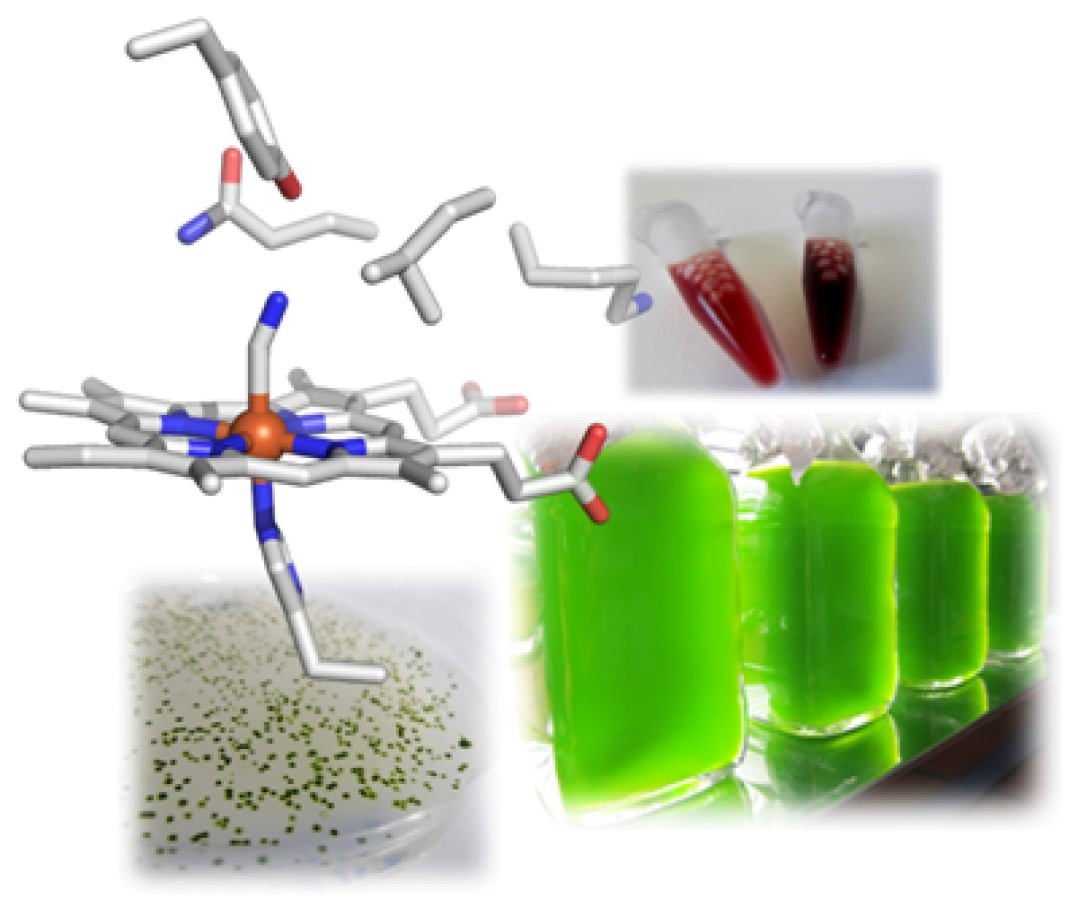
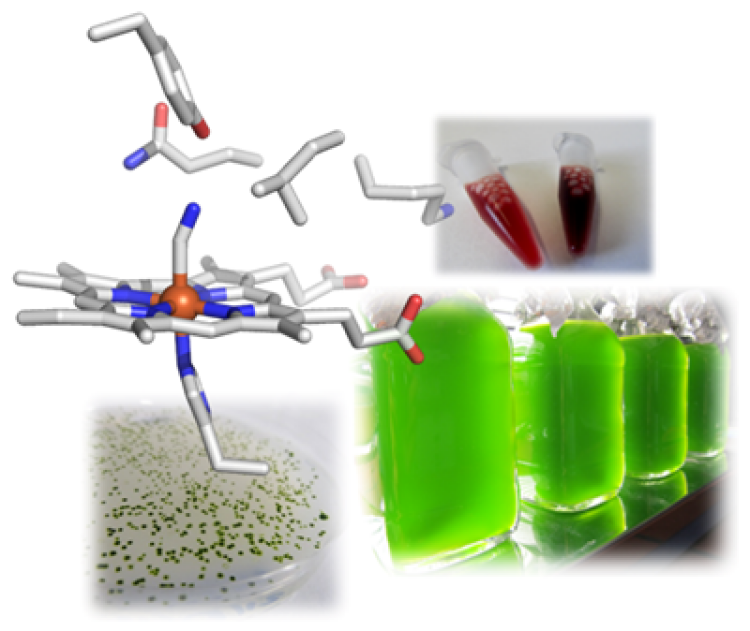
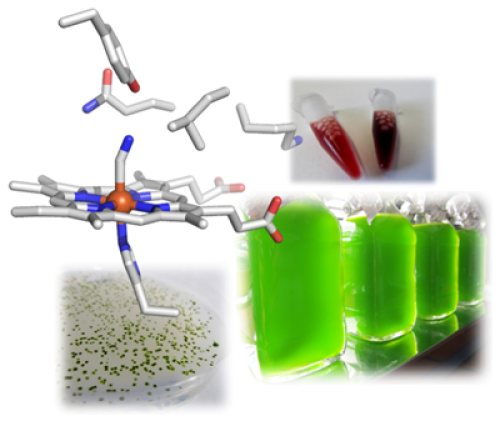
Hier wenden wir z.B. Transcriptomics an, um Gene zu identifizieren, die besonders stark auf Hypoxie reagieren. In den Daten haben wir Gene bzw. die kodierten Proteine entdeckt, die vermutlich an einer Stickstoffmonoxid- (NO-) basierten Signaltransduktion beteiligt sind, wie etwa eine so genannte NO-sensitive Guanylatcyclase. Dieses Enzym ist sehr ähnlich zu den entsprechenden Enzymen in Tieren und Menschen, die durch NO aktiviert werden und dann den sekundären Botenstoff cGMP bilden. Da dieser Botenstoff ubiquitär vorkommt, interessiert uns ebenfalls, welche Zielproteine durch cGMP direkt oder indirekt reguliert werden. Hier vermuten wir Verbindungen zum Stärke-Stoffwechsel von C. reinhardtii und charakterisieren daher einzelne Enzyme des Stärke-Aufbaus und -Abbaus.
Weiterhin sind wir während dieser Forschungsarbeiten auf eine bestimmte Klasse an Hämoglobin-artigen Proteinen in der Mikroalge aufmerksam geworden. Anders als allgemein bekannt besitzen fast alle – auch einzellige und prokaryotische – Organismen Hämoglobin-Formen und nutzen diese für viele andere Prozesse als den bekannten Transport von O2, unter anderem für die katalytische Bildung oder die Entgiftung von NO. Die Mikroalge C. reinhardtii besitzt zwölf dieser Proteine, die sehr divers sind. Uns fasziniert die Fragestellung, warum ein Einzeller solch eine Vielfalt an Hämoglobinen benötigt und erforschen die Proteine hinsichtlich der von ihnen katalysierten Reaktionen mit einem Fokus auf einer möglichen Beteiligung am NO-Stoffwechsel. Letztendlich möchten wir herausfinden, welche Rolle sie in der Alge spielen. Hier haben wir seit der Entdeckung des CRISPR/Cas-Systems als Gen-Editierungswerkzeug erstmal die Möglichkeit, Gene in C. reinhardtii gezielt auszuschalten oder auch nur einzelne Mutationen einzufügen, um so die zellulären Funktionen unserer Zielproteine zu entschlüsseln.
Copyright © pbt 2026
Letzte Änderung: 22. Jan. 2026
{kind=link}
{kind=link}
{kind=link}