![]()
![]()
![]()
![]()
Senior Lecturer
Room:
ND 2/136
Phone: +49 234 32 -
23830
Academic Council
ORCID-iD: 0000-0001-8879-3348
E-Mail

Pro- and eukaryotic photosynthetically active microorganisms play an enormously important role for the ecosystem Earth. They are responsible for a large part of the photosynthetic formation of O2 and biomass and, in parallel, the assimilation of CO2, thus ensuring the survival of non-photosynthetic organisms, including humans. In their natural environment, all organisms experience stress frequently, such as the deficiency or excess of a nutrient. Because of the highly energetic processes involved in the light reactions, photosynthetic organisms must be particularly capable of responding to stress with appropriate defensive measures. Photosynthetic H2 production by microalgae represents such a stress response and serves to dissipate excess light energy to prevent oxidative damage to the cell. It is frequently induced upon nutrient deficiency, which is often accompanied by the establishment of O2-limited (hypoxic) conditions in algal cultures. Our early research has helped to understand the very elaborate fermentation metabolism of the microalga Chlamydomonas reinhardtii, which in its complexity is unusual for a eukaryote and of which H2 production is a part. In addition to providing fascinating insights into the ecology of this alga, which in turn is crucial for functional ecosystems, the study of the acclimation of microalgae to O2 deficiency is also biotechnologically relevant. On the one hand, some of the fermentation products (e.g. ethanol or lactic acid) represent industrially relevant substances, and on the other hand, algal cultures often experience hypoxic conditions in economically established cultivation tanks. In addition to charaterizing metabolism under these conditions, we are also trying to understand how the alga 'notices' that O2 is missing in the first place, and how it relays this information in the cell to activate the appropriate responses – often at the level of gene expression.
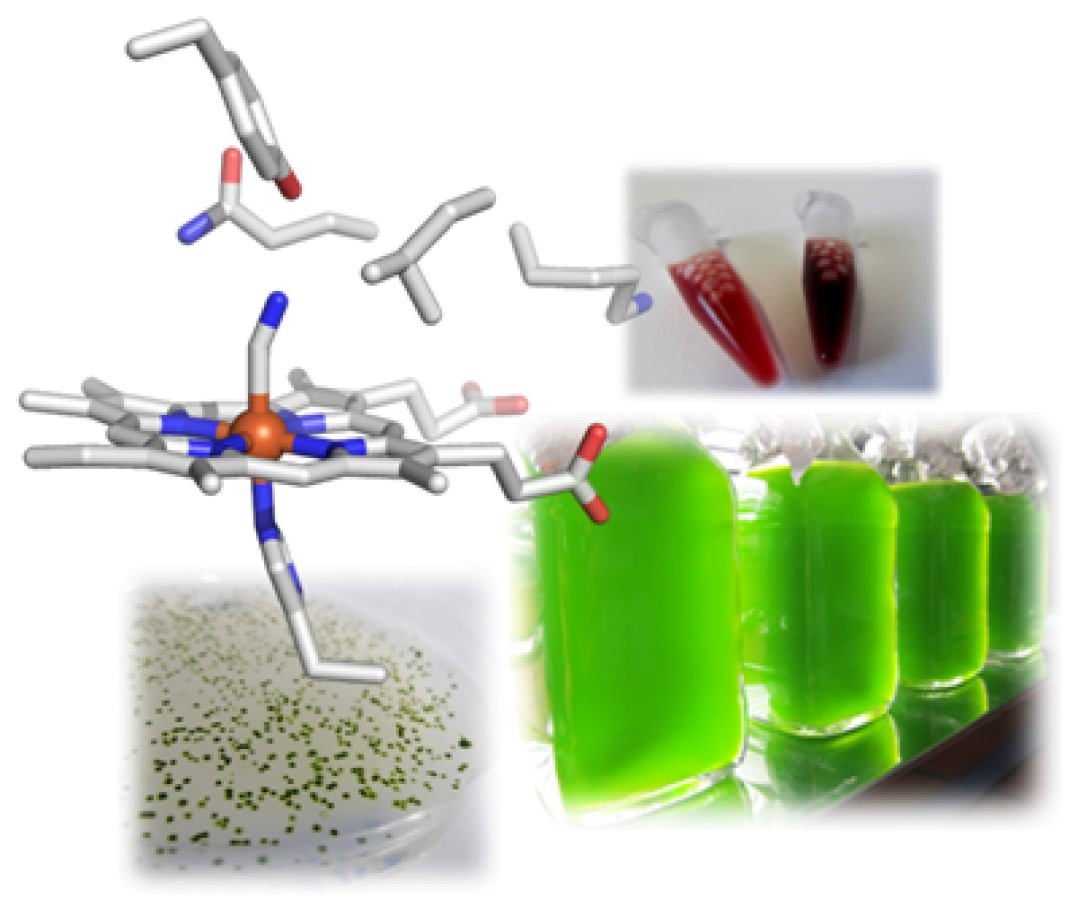
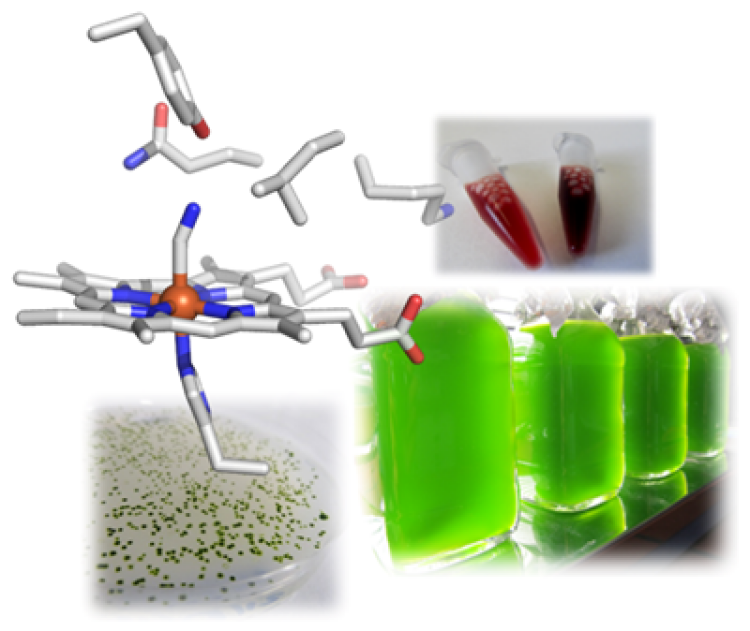
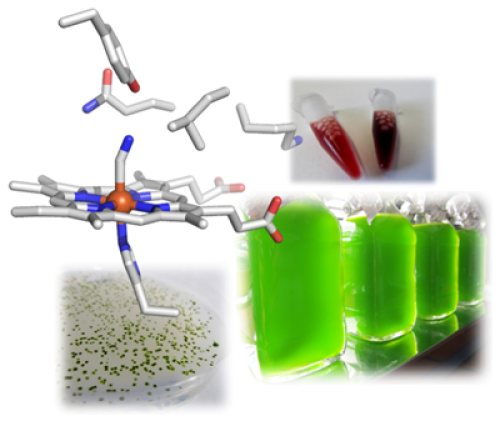
To this end, we apply transcriptomics to identify genes that respond particularly strongly to hypoxia. In the data sets, we have discovered genes and the encoded proteins, respectively, that are thought to be involved in nitric oxide (NO)-based signal transduction, such as a so-called NO-sensitive guanylate cyclase. This enzyme is very similar to the corresponding enzymes in animals and humans that are activated by NO and then form the secondary messenger cGMP. Since this messenger is ubiquitous, we also seek to identify target proteins that are directly or indirectly regulated by cGMP. Here, we suspect connections to the starch metabolism of C. reinhardtii and therefore characterize individual enzymes of starch breakdown and degradation.
Furthermore, in the course of this research we became aware of a particular class of hemoglobin-like proteins in the microalga. Contrary to common knowledge, almost all organisms – including unicellular and prokaryotic ones – possess hemoglobins and use them for many processes other than the well-known transport of O2, including catalytic formation or detoxification of NO. The microalga C. reinhardtii has twelve of these proteins, whose primary structures are very diverse. We are intrigued by the question of why a unicellular organism requires such a diversity of hemoglobins and are exploring the proteins in terms of the reactions they catalyze with a focus on possible involvement in NO metabolism. Ultimately, we want to understand the role they play in the alga. Since the discovery of the CRISPR/Cas system as a gene editing tool, we have for the first time the possibility to selectively knock out genes in C. reinhardtii or even to insert single mutations in order to decipher the cellular functions of our target proteins.
Copyright © pbt 2026
Last update: Jan 22, 2026
{kind=link}
{kind=link}
{kind=link}